Have you ever experienced museum fatigue? I thought that I made up this term to describe my own experiences, but upon performing a quick Google search, I discovered that this is actually a phenomenon first described in 1916 (Gilman, 1916).

Interior of the Musée d’Orsay (Image from TripSavy.com)
Going to a museum may seem like a passive process, but to me, it is actually quite a bit of work!
Navigating large crowds and carrying a heavy backpack for several hours is enough to wear me out. But even more so, interpreting piece after piece of artwork—each of which leaves a lot of room for interpretation—is a laborious effort leading to mental exhaustion. Though it is uncomfortable, I think that this is the way it should be. If you don’t experience some fatigue, are you fully engaged with and appreciating the art?

Exterior of Musée d’Orsay (Image from SortiraParis.com)
One particular French artist I have learned about in class is Paul Cézanne, and he seems to have been an especially avid proponent of museum fatigue; although his works were rejected from museums during his lifetime, it seems as if he were intentionally inducing this exhaustion. In the Post-Impressionistic style (abandoning the detailed, picture-perfect landscapes characteristic of Realism), Cézanne produced blurry, unfinished images in order to accentuate the mind’s interpretation process. Leaving blank spots peeking through the blobs of color is a technique called nonfinito, and it’s a bit like trailing off in the middle of a sentence—a visual ellipsis. In this way, the viewer’s interpretation is unique to the way the mind fills in the gaps at that particular moment, influenced by all of the emotions and experiences one brings to the table.
It turns out that this reflects how the brain works when interpreting all visual stimuli: even looking at the same things twice may trigger different responses from neurons dedicated to processing visual information (Jeon et al., 2018).
First, let’s start with some background information about vision and how our

The occipital lobe, shown in yellow (Image from The Science of Pscychotherapy.com)
brains process signals coming from our eyes.
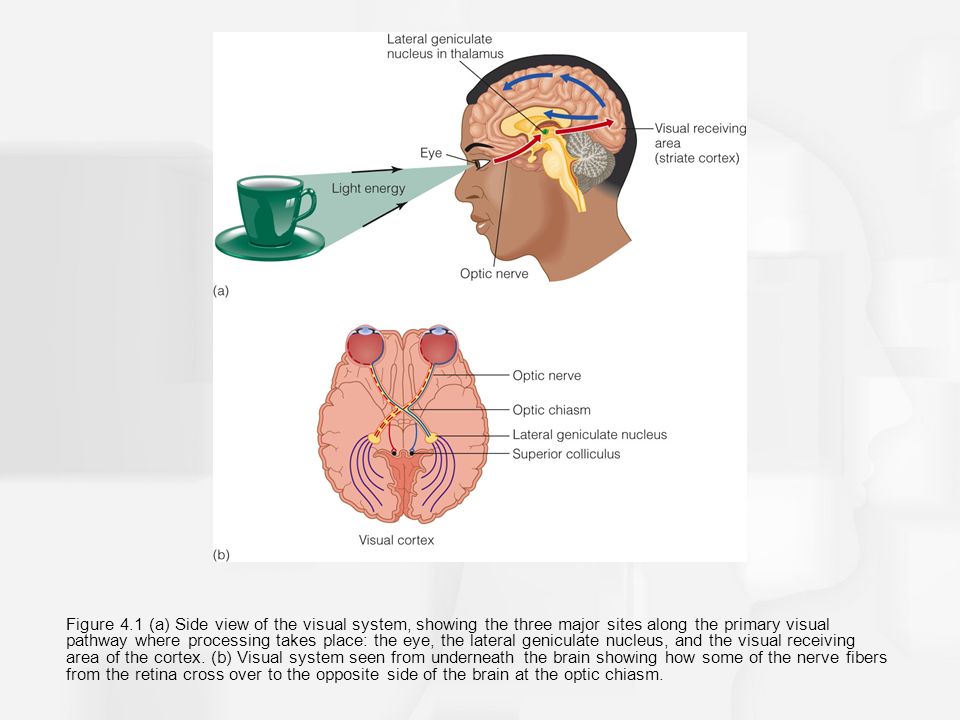
Light enters the eye and reaches the retina at the very back. There, it stimulates light-responsive cells called photoreceptors (rods and cones). Signals from all these cells go through the optic nerve, the optic tract, a structure called the thalamus, and eventually reach the part of the brain that deals with visual information. This area is called the occipital lobe, and the section that is first to receive these signals is called the primary visual cortex, or V1. Here, there are cells that have been shown to respond to basic details of a scene like the width and orientation of lines (Gawne, 2015). Each cell is “tuned” to respond best to a certain width and a certain orientation, and logically, this is called neuronal tuning (Butts and Goldman, 2006). The conditions determining the responsivity of the neurons get more and more complex as the signals are processed (Tsunoda et al., 2001).

The perception of visual information (Image from Slideplayer.com)
As one views the same image, it would make sense that the same neurons respond each time. But, this is not exactly the case: In one experiment by Jeon et al. 2018 in the journal Nature, researchers found that the same neurons aren’t reliably activated by the same stimuli.
In the study, the researchers showed mice lines of different orientations and widths. Using a technique called two-photon calcium imaging, they looked at the activity of neurons in the V1 (Jeon et al., 2018). This technique involves installing an apparatus on the head of a mouse. Based on the movement of fluorescing ions, it lets us see what neurons are active as the mouse is awake and interacting with the world (Mitani and Komiyama, 2018).

Some of the images shown to mice in the Jeon et al. (2018) experiment (Image from the journal Nature)
Tracking around 300 neurons, the researchers determined the qualities of the image (such as the angle and the width of the lines) for which a neuron was most likely to respond. Then, performing the test one week later and again two weeks later, they compared the preferences of the neurons. While the majority of individual qualities were relatively stable over time, the researchers found that fewer than half of the neurons had exactly all of the same preferences as before.
What does this all mean? In the past it has been shown that the visual cortex is highly plastic, or able to rearrange and reorganize its connections based on new information (Hofer et al., 2009). However, these results provide even more insight into how our visual systems adapt and change: some parts can remain stable while others change their responsivity in order to incorporate new information, altering our perception of the world around us.
So, our perception of static scenes is actually not static at all; it is being altered constantly! That boulangerie we pass on the way to class is not perceived by our brains in exactly the same manner every day.

Portrait of a Woman by Paul Cezanne (Image from the Metropolitan Museum of Art)
That leads me to wonder: especially when looking at one of Cézanne’s paintings—since he left so much for the viewer’s mind to fill in—do we ever experience the same thing twice? This may very well be the most intriguing thing about his work, making it both timeless and malleable. A perfect excuse to visit the Musée d’Orsay just one more time. The unfortunate result is only that this “museum fatigue” may become an increasingly common affliction. However, it’s likely already a common experience for all the museum-goers of the world, and I’m not afraid. It certainly won’t deter me from absorbing all of the Post-Impressionism art I can while I’m here!
References:
Butts, D.A., Goldman, M.S. (2006). Tuning curves, neuronal variability, and sensory coding. PLOS Biology. 4:92. doi: 10.1371/journal.pbio.0040092.
Gawne, T. (2015). The responses of V1 cortical neurons to flashed presentations of orthogonal single lines and edges. Journal of Neurophysiology. 113:2676-2681. doi: 10.1152/jn.00940.2014
Gilman, B. I. (1916). Museum Fatigue. The Scientific Monthly. 2:62–74.
Hofer, S. B., Mrsic-Flogel, T. D., Bonhoefer, T. & Hubener, M. (2009). Experience leaves a lasting structural trace in cortical circuits. Nature. 457:313–317.
Jeon, B. B., Swain, A.D., Good, J. T., Chase, S. M., Kuhlman, S.J. (2018). Feature selectivity is stable in primary visual cortex across a range of spatial frequencies. Nature. 8:15288. doi:10.1038/s41598-018-33633-2.
Mitani, A., Komiyama, T. (2018). Real-time processing of two-photon calcium imaging data including lateral motion artifact correction. Frontiers in Neuroinformatics. 12:98. doi: 10.3389/fninf.2018.00098
Tsunoda, K., Yamane, Y., Nishizaki, M., Tanifuji, M. (2001). Complex objects are represented in macaque inferotemporal cortex by the combination of feature columns. Nature Neuroscience. 4:832-838. doi: 10.1038/90547.
Image links:
https://www.sortiraparis.com/images/55/1467/108625-jeu-de-piste-gratuit-au-musee-d-orsay.jpg
https://collectionapi.metmuseum.org/api/collection/v1/iiif/656892/1571903/restricted
Hyperlinked videos and sites:
https://www.youtube.com/watch?v=fZDAwXh54is
https://goo.gl/maps/GM2ZURPTLpia3V7b9



:max_bytes(150000):strip_icc()/8414359797_20e28f27f2_o-56a403d25f9b58b7d0d4f0e6.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}