
Contributed by Alexandria Albert and Gavon Broomfield
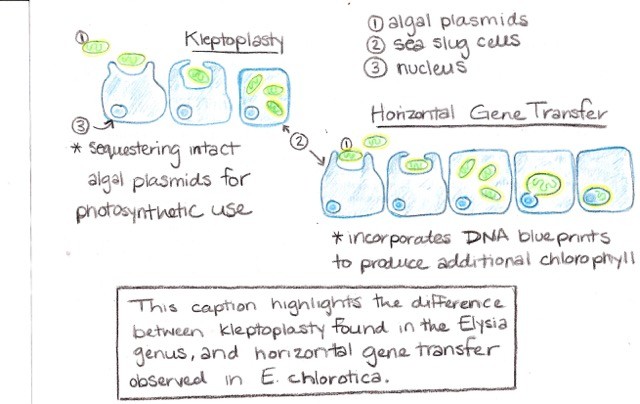
Bright green sea slugs that behave like plants! Sea slugs are a diverse family of marine gastropod mollusks characterized by their soft bodies and lack of external shell. Approximately 2,300 species have been documented, all with different physical colorations that allow them to better interact with other organisms and underwater conditions. Sacoglossan sea slugs have mastered the art of kleptoplasty by extracting chloroplasts from various algal food sources and preserving them in digestive tissue, thus creating the kleptoplast. Exploring this symbiotic interaction has provided insight into multiple evolutionary processes. For example horizontal gene transfer, the transfer of genes from one species to another, in this case from the algal nucleus to sea slug cells, has facilitated the long-term use of the kleptoplasts (Cruz et al. 2013). There are still questions about the maintenance of kleptoplasts living in animal tissue, but benefits from this form of energy production have been documented.

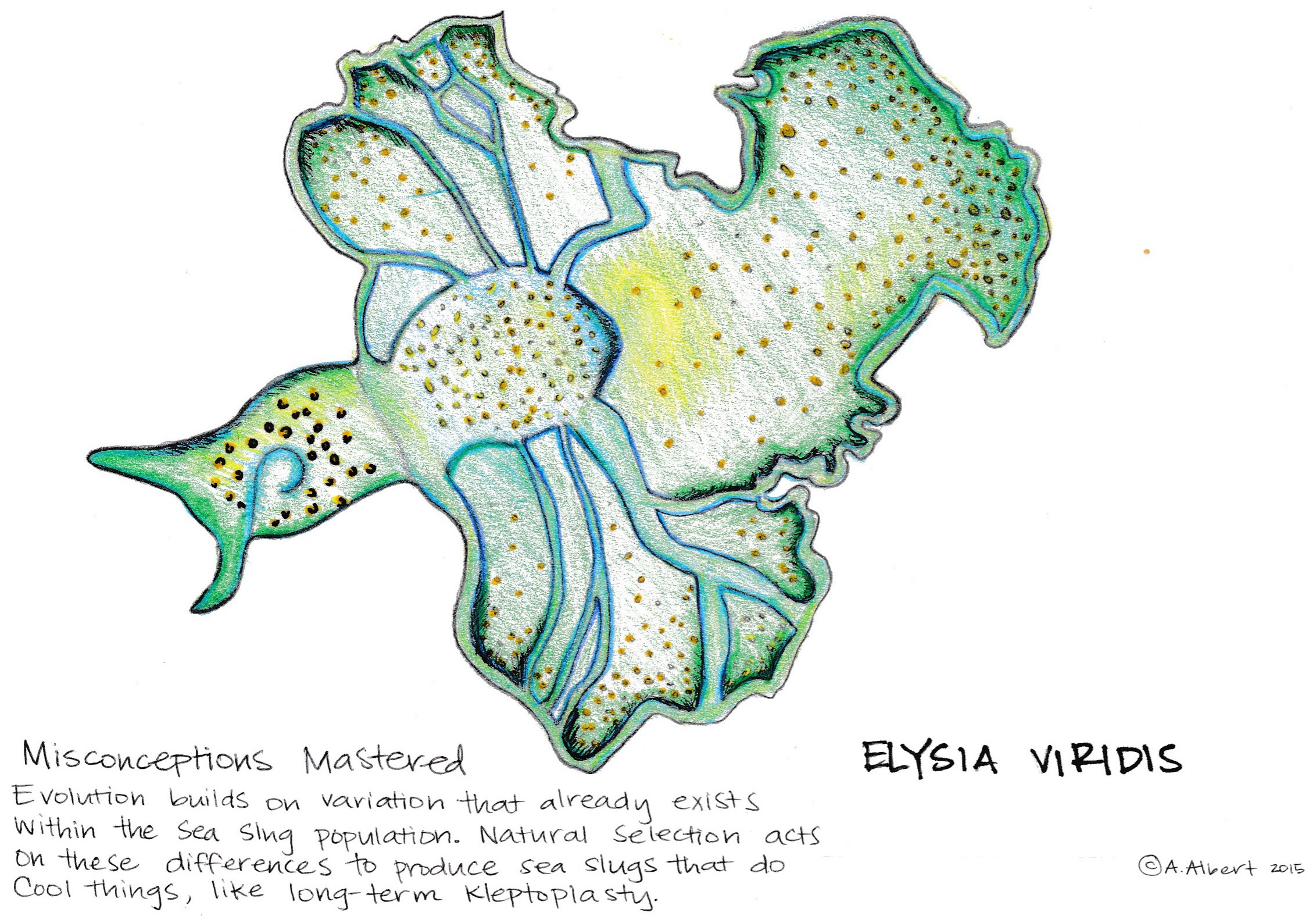
So how is it possible that sea slugs have chloroplasts? Aren’t chloroplasts only in plants? The key to photosynthetic capable sea slugs is symbiosis. Algal nuclear genes in the sea slug digestive cells encode for chlorophyll synthesis, giving slugs green coloring, and chloroplast proteins, which later become incorporated into the slug’s DNA to get passed onto offspring. For some species of Sacoglossa, these internal or endosymbiotic chloroplasts can be maintained long-term if the slug possesses the nuclear DNA required for photosynthesis. For other species, continual feeding on algae is necessary for long-term, sustained kleptoplastic ability. Cool, right? Since the chloroplast is not native to the sea slug, important behavioral, morphological, and biochemical adaptations have evolved to maintain this symbiosis and kleptoplasty (Schwartz, Curtis, and Pierce 2014, Schmitt, Valerie, et al. 2014).
So how do these photosynthetic sea slugs use these chloroplasts? Sea slugs adjust their parapodial lobes, lateral fleshy protrusions on their bodies used for movement, to manage light harvesting. When the parapodial lobes are extended, chloroplasts are exposed to direct sunlight which is then used as an energy source, a process known as phototrophy, as seen in plants. When doing this, sea slugs often resemble leaves. This leaf-like appearance aids in camouflage and avoidance of ocean-floor predators like crab, lobster, and fish (Schmitt and Wägele 2011). This unique behavioral adaptation has evolved to retain endosymbiotic chloroplasts.
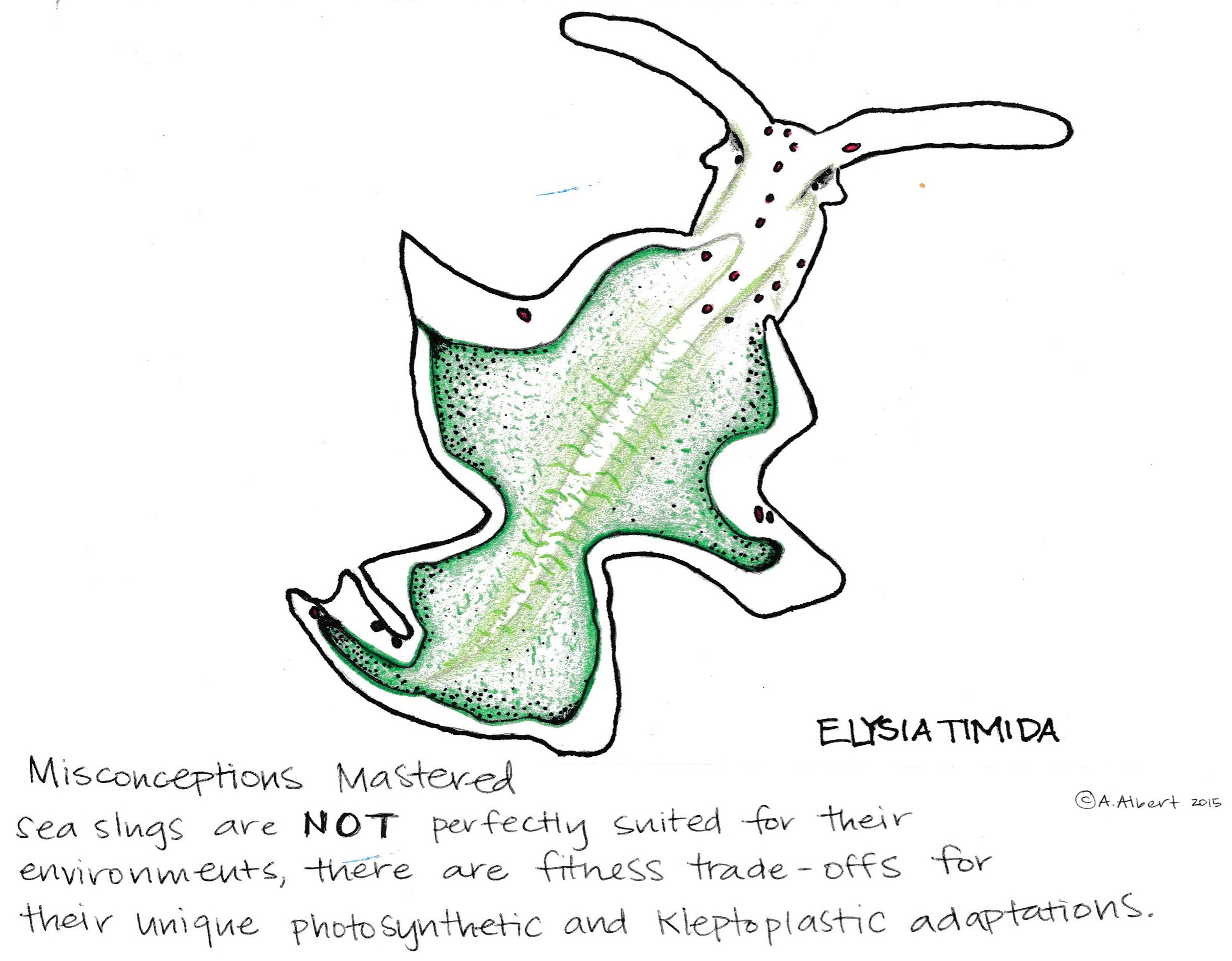
Does this really work? Studies have shown high levels of fitness benefits to kleptoplasty in Sacoglossa when measuring growth efficiency with trade-offs dependent upon algae diet and light exposure (Baumgartner, Pavia, and Toth 2015). This adaptation has some limitations and may depend upon sunlight exposure and the species of algae. Too much sun exposure could cause photo-oxidative stress on kleptoplasts and decrease the rate of energy production over time (Serôdio, João et al. 2014).
For more information about the photosynthetic qualities of sea slugs, consult these sources:
- Baumgartner, Finn A., Henrik Pavia, and Gunilla B. Toth. “Acquired Phototrophy through Retention of Functional Chloroplasts Increases Growth Efficiency of the Sea Slug Elysia Viridis.” Ed. Erik Sotka. PLoS ONE 10.4 (2015): e0120874. PMC. Web. 6 Nov. 2015.
- Goodheart, J. A., Bazinet, A. L., Collins, A. G., & Cummings, M. P. (2015). Relationships within Cladobranchia (Gastropoda: Nudibranchia) based on RNA-Seq data: an initial investigation. Royal Society Open Science, 2(9), 150196. http://doi.org/10.1098/rsos.150196
- Schmitt, Valerie, and Heike Waegele. “Behavioral adaptations in relation to long-term retention of endosymbiotic chloroplasts in the sea slug Elysia timida (Opisthobranchia, Sacoglossa).” Thalassas 27.2 (2011): 226-238.
- Schwartz, Julie A., Nicholas E. Curtis, and Sidney K. Pierce. “FISH labeling reveals a horizontally transferred algal (Vaucheria litorea) nuclear gene on a sea slug (Elysia chlorotica) chromosome.” The Biological Bulletin 227.3 (2014): 300-312.
- Schmitt, Valerie, et al. “Chloroplast incorporation and long-term photosynthetic performance through the life cycle in laboratory cultures of Elysia timida (Sacoglossa, Heterobranchia).” Frontiers in zoology 11.1 (2014): 5.
- Serôdio, João et al. “Photophysiology of Kleptoplasts: Photosynthetic Use of Light by Chloroplasts Living in Animal Cells.” Philosophical Transactions of the Royal Society B: Biological Sciences 369.1640 (2014): 20130242. PMC. Web. 6 Nov. 2015.






 In fact, these two rattlesnakes are very closely related, but the populations that live in the mountains have a distinctly different venom than those that live in the desert.
In fact, these two rattlesnakes are very closely related, but the populations that live in the mountains have a distinctly different venom than those that live in the desert.







